The diaphragm develops from four embryonic structures: the septum transversum, the pleuroperitoneal membranes, the dorsal mesentery of the esophagus, and the body cavity walls, each of which either becomes totally incorporated into the diaphragm or merely contributes part of itself to the diaphragm (Moore, 1982). Before discussing these strucutres directly, however, it will be helpful to set the scene for diaphragm development by outlining the development of the major body cavity subcavitites and tissues as detailed in Moore (1982).
The body cavity, or coelom, begins as two spaces in the lateral and cardiogenic mesoderm, lateral to the somites. By the fourth week, the two spaces have fused to form one horseshoe-shaped cavity. The bend at the anterior of the cavity will become the pericardial cavity (surrounding the heart), and the two "limbs" (lateral parts) will become the pleural and peritoneal cavities. The pleural cavity is the thoracic cavity, surrounding the lungs. The peritoneal cavity contains the primordium of the abdominal and pelvic viscera; its parietal wall is lined by mesothelium, which is future peritoneum and is derived from somatic mesoderm. The visceral wall is covered by mesothelium derived from splanchnic mesoderm. As the embryonic folding occurs, the two "limbs" come together in the ventral part of the embryo. The ventral mesentery in this area degenerates, leaving the large peritoneal cavity. The pericardioperitoneal canals, positioned lateral to the future esophagus and dorsal to the septum transversum, separate the peritoneal and pericardial cavities. The pleural cavities form when partitions separate the pericardial and pleural cavities and the pleuritoneal and pleural cavities. The two partitions that gradually separate the pleural cavity from the peritoneal cavity are the pleuroperitoneal membranes. They form as the lungs and pleural cavities expand into the body walls.
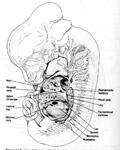
The first strucuture to become part of the diaphragm is the septum transversum, a mesodermal strucutre that forms the central tendon of the diaphragm. The septum transversum is identifiable at three weeks of gestation as a mass of mesoderm just posterior to the pericardial cavity, which contains the heart. In the fourth week, after the head fold forms, the septum thickens to become a partial diaphragm between the pericardial and peritoneal (abdominal) cavities. It fuses with the mesenchyme ventral to the esophagus and with the pleuroperitoneal membranes, thus becoming the central tendon of the diaphragm.
The pleuroperitoneal membranes fuse with the dorsal mesentery of the esophagus and with the dorsal portion of the septum transversum to complete the partition between the pleural and peritoneal cavities. The membranes form the primitive diaphragm but end up as only a small portion of the fully developed diaphragm.
The dorsal mesentery of the esophagus fuses with the two structures septum transversum and the pleuroperitoneal membranes to form the middle of the diaphragm. Other muscle fibers that grow into the dorsal mesentery of the esophagus will develop into the crura of the fully developed diaphragm. Crurae are pairs of diverging bundles of muscle.
During weeks 9 through 12, the body-wall tissue splits into two layers as a result of the enlargement of the lungs and the pleural cavities. The outer layer will form the true body wall, and the parts of the inner layer will form the peripheral parts of the diaphragm. These parts will be between the body wall and the structures from the pleuroperitoneal membranes. As the pleural cavities extend further in to the body walls, the costodiaphragmatic recesses form, which become the costa. This extension forms the dome-shape of the adult diaphragm. Postpartum, the costodiaphragmatic recesses expand and contract in size as the lungs expand and contract during breathing.
In a frequently-cited study DeTroyer et al. (1981) reported that the diaphragm consists of two distinct muscles that each have different actions on the rib cage. This makes sense in terms of the embryonic origins of the diaphragm, since it forms as multiple muscle bundles come together, rather than as a single sheet of tissue that separates into multiple structures. As reported above, myoblasts from the lateral body walls become the costal parts, and the dorsal mesentery of the esophagus forms the crural parts. These two muscles have different innervation origins as well. When the costal muscle of dogs was electrically stimulated, lung volume and abdominal pressure increased, and there was an outward displacement of the abdomen and the lower rib cage. Electrical stimulation of the crural muscle had similar effects except there was no effect on the rib cage dimension. The crural muscle, which is attached to the lumbar vertebrae but not attached to the rib cage, functions mainly to exert abdominal pressure.